Cовременные технологии физического развития и воспитания детей дошкольного возраста.
I. История возникновения и развития налогов и налогообложения в дореволюционной России
II. История развития налогов и налогообложения в СССР. Становление налоговой системы современной России
II. От распространения образования и развития трудолюбия и бережливости.
II. ПРОБЛЕМЫ И ОСНОВНЫЕ ФАКТОРЫ РАЗВИТИЯ ТОПЛИВНО-ЭНЕРГЕТИЧЕСКОГО КОМПЛЕКСА
T Этапы развития средств вычислительной техники.
V. Размножение и циклы развития водорослей.
V1. Предмет психологии. Методы психологии. История развития научной психологии
VII. РЕГИОНАЛЬНЫЕ ОСОБЕННОСТИ РАЗВИТИЯ ЭНЕРГЕТИЧЕСКОГО СЕКТОРА
VII. Третий пояс педагогических исследований изучение развития человека
VIII. Методы исследования системы обучения и развития, как научная и конструктивная проблема
Windows. История развития.
Классификация растений, их характеристика и жизненные циклы
Мхи или моховидные – отдел высших растений, объеденных в классы: печёночные мхи (6 000–8 000 видов), антоцеротовые мхи (100–200 видов) и листостебельные мхи (10 000 видов). Мхи это, как правило, мелкие растения, длина которых редко превышает 50 мм. Отличаются от других высших растений тем, что в их жизненном цикле взрослые растения представлены разнополыми гаметофитами, а спорофит развивается на женском гаметофите. Размножаются бесполым путём – спорами и вегетативно частями таллома.
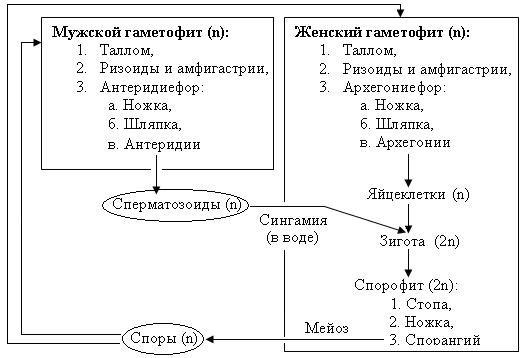
В цикле развития взрослые (многолетние) растения маршанции полиморфной представлены разнополыми (двудомными) мужскими и женскими гаметофитами. Вегетативное тело растения – пластинчатый таллом, имеет вид мясистого лопастного, дихотомически разветвленного слоевища длиной до 10-12 см и шириной до 3 см, образованного множеством клеточных слоев (около 30). Специализированных проводящих тканей таллом не имеет. На нижнем эпидермисе образуются два типа выростов:
– ризоиды – бесцветные простые и язычковые одноклеточные выросты, которые выполняют функцию закрепления растения в субстрате. Воду маршанция способна поглощать всем телом,
– амфигастрии – окрашенные (темно-фиолетовые) однослойные, многоклеточные чешуи (редуцированные «листья» – филоиды), которые способны долго удерживать воду во время засухи.
Различить мужской и женский гаметофиты возможно во время развития на поверхности талломов специальных органов (подставок для гаметангиев) – мужских антеридиефоров и женских архегониефоров.
Антеридиефор представляет собой ножку с расположенным сверху восьмиугольным диском (шапкой). С верхней стороны шапки в антеридиальных камерах находятся мужские гаметангии – антеридии. В антеридиях образуются мужские двужгутиковые гаметы – сперматозоиды.
Архегониефоры развиваются на женских гаметофитах в виде девятилучевого щитка (звездочки) на ножке. Во время развития архегониефора, архегонии (женские гаметангии) закладываются с морфологически верхней стороны подставки. В силу неравномерного роста обеих сторон ложа подставки (щитка) они смещаются между лучами щитка на его нижнюю сторону, где располагаются группами. Вокруг каждой группы архегониев образуется специальная защитная обертка. Архегонии представляют собой кувшиноподобные камеры, направленные горловиной вниз, в брюшке которых образуется по одной яйцеклетке.
Половой процесс в цикле развития маршанции происходит посредством воды преимущественно во время дождя. Капли дождя выбивают из антеридиев зрелые сперматозоиды и вместе с ними отражаются на женские шапки архегониефоров. Сбегая между лучами женских подставок, капли воды вместе со сперматозоидами зависают в месте расположения архегониев, покрывая их. По воде мужские гаметы проникают через шейку архегония в брюшко – калиптру, где оплодотворяют яйцеклетку, образуя зиготу (первую диплоидную клетку спорофита). После оплодотворения вокруг каждого архегония из его ножки начинает образовываться индивидульная чашевидная защитная обертка – 4-5 лопастной псевдоперианций. В это время зигота делится митозом, формируя зародыш спорофита, который прикрепляется к материнскому телу (щитку архегониефора) присоской – гаусторией, и получает оттуда питание. С увеличением размеров зародыша калиптра (брюшко архегония) растягивается (увеличивается в размерах).
Взрослый спорофит, развивающийся из зародыша, включает в себя: стопу (присоску) – закрепляющую часть, ножку и спорангий (коробочку спорогона). В процессе своего роста спорофит разрывает калиптру и выносит спорангий наружу. В спорангии формируются диплоидные клетки спорогенной ткани, которые получают питание через удлиненные клетки со спирально утолщенными клеточными стенками – элатеры. В результате разделения спорогенных клеток мейозом образуются гаплоидные гетероталличные споры. При созревании спор элатеры высыхают и скручиваются спирально, разрыхляя массу спор в спорангии. Однослойная коробочка спорангия при созревании спор легко раскрывается восемью загнутыми назад зубцами, а пружинки элатер способствуют равномерному рассеиванию спор. Гетероталличные (разнополые) споры маршанции в благоприятных условиях прорастают в протонемы (передростки) – небольшие беспигментные нитчатые выросты, с верхушечной клетки которых образуются талломы соответственно мужского и женского гаметофита нового поколения.
Вегетативное размножение маршанции происходит с помощью выводковых почек, которые формируются в специальных выростах – выводковых корзинках, на верхней стороне как женских, так и мужских талломов. В результате попадания воды на таллом выводковые почки вымываются (распрыскиваются) и прорастают на почве в новые талломы соответствующего пола. Маршанция распространена в увлажненных местах на берегах водоемов, под покровом леса.
Рис. 30. Схема жизненного цикла мха маршанция
Отдел моховидные, их строение и происхождение
Моховидные (Bryophta) — не самая древняя, но самая примитивная группа современных наземных растений, сохранившая много черт сходства с водорослями. Они появились в девоне, около 370-400 млн лет т.н. Единого мнения о происхождении моховидных нет. Существует несколько гипотез их происхождения. Некоторые ученые полагают, что моховидные — это редуцированные сосудистые растения; другие считают, что и моховидные, и сосудистые растения произошли от единого наземного предка; наконец, наиболее распространенной и общепринятой является гипотеза, согласно которой моховидные и сосудистые растения произошли независимо друг от друга от различных видов зеленых водорослей.
Таким образом, по мнению большинства ученых, моховидные и сосудистые растения представляют собой две различные эволюционные линии развития высших растений и имеют отдаленного общего предка среди зеленых водорослей.
Родство зеленых водорослей и наземных растений (как моховидных, так и сосудистых) подтверждается в первую очередь одинаковым набором фотосинтетических пигментов: основным фотосинтетическим пигментом у них служит хлорофилл а, вспомогательными пигментами — хлорофилл в и каротиноиды (включая ксантофиллы), а также наличием целлюлозы в клеточной стенке и накоплением питательных веществ в пластидах, а не непосредственно в цитоплазме, что характерно для других отделов водорослей.
Особенности строения и развития моховидных
Необходимый элемент строения каждого наземного растения покровные ткани, защищающие растение от высыхания. Они сеть у всех наземных растений, однако у моховидных покровные ткани развиты хуже, чем у сосудистых растений. Часто они менее специализированы и содержат хлоропласты, т.е. выполняют не только защитную, но и фотосинтетическую функцию. У многих моховидных, в частности у торфяною мха сфагнума, листья состоят только из одного слоя клеток и, естественно, дифференцированной покровной ткани не имеют.
У большинства моховидных покровные ткани не имеют воскового налета — кутикулы, препятствующего испарению воды, и вследствие этого плохо защищают растение от высыхания.
Наличие покровных тканей предполагает и наличие устьиц отверстий для осуществления газообмена. Однако у большинства моховидных устьица устроены и функционируют иначе, чем у сосудистых растений. Они не могут тонко регулировать газообмен, постоянно открываясь и закрываясь. Устьица моховидных постоянно открыты и закрываются только тогда, когда растение высыхает.
У моховидных, так же как у водорослей, нет корней. Их заменяют выросты покровной ткани — ризоиды. Ризоиды мхов, так же как ризоиды водорослей, удерживают растение в грунте, но воду всасывают плохо. Поэтому моховидные, так же как и водоросли, поглощают воду всей поверхностью тела (в первую очередь листьями).
У моховидных отсутствуют или плохо развиты проводящие ткани.
У большинства моховидных проводящих тканей нет. У некоторых они представлены только тяжем гидроидов — мертвых клеток, напоминающих сосуды ксилемы, и только у самых сложно устроенных моховидных (зеленые мхи, или брииды) внутри «стебля» фотосинтезирующего поколения гаметофита проходит тяж гидроидов, окруженный клетками лептоидов, по строению и функциям напоминающих ксилему и флоэму сосудистых растений.
У большинства моховидных отсутствуют специализированные механические ткани, а гидроиды, напоминающие сосуды ксилемы, нс имеют специальных утолщений, которые помогают выполнять опорную функцию клеткам ксилемы сосудистых растений.
Вследствие того что у моховидных плохо развиты проводящая и механическая ткани, они никогда не вырастают высокими — у большинства из них высота не превышает 20 см.
Но самой удивительной особенностью представителей этого отдела, резко отличающей их от всех остальных наземных растений, является то, что в их жизненном цикле доминирует не диплоидное поколение — спорофит (2n), а гаплоидное поколение — гаметофит (n).
Именно гаплоидный гаметофит осуществляет у моховидных основную функцию растения — фотосинтез.
У большинства моховидных гаметофит имеет форму стебля с листьями, однако названия «стебель» и «лист» в данном случае условны, поскольку эти органы у моховидных и сосудистых растений не гомологичны, а лишь аналогичны. Если у сосудистых растений стебель и лист — это элементы диплоидного поколения — спорофита, то у моховидных они — элементы гаплоидного поколения — гаметофита.
Фотосинтезирующий гаметофит мхов снабжает питательными веществами как себя, так и диплоидное поколение — спорофит. Спорофиту моховидных редуцирован и обычно паразитирует на гаметофите.
Особенности размножения и развития моховидных особенно хорошо видны в жизненном цикле печеночного мха — маршанции многообразной (Marchantia polymorpha).
Жизненный цикл маршанции
В лесу, на влажной почве, можно встретить растения, которые выглядят необычно и загадочно, как инопланетяне. Это растения печеночного мха — маршанции. (Marchantia polymorpha)
В отличие от зеленых мхов, гаметофит маршанции — не стебель с листьями, а простая, дихотомически ветвящаяся зеленая пластинка, похожая на извлеченную из воды пластинчатую водоросль. Такое просто устроенное тело у моховидных, так же как и у водорослей, называется слоевищем, или талломом.
Рассмотрим цикл развития маршанции, начиная с прорастания спор (рис. 1).
У маршанции (так же как у всех водорослей и наземных растений) споры гаплоидны (n), т.е. несут одинарный набор хромосом. Споры у маршанции разные: одни из них мужские (микроспоры), а другие — женские (мегаспоры). Из спор вырастают соответственно мужской и женский гаметофиты. Гаметофиты, сформировавшиеся путем деления исходных гаплоидных спор (n), тоже гаплоидны (n). Мужской и женский гаметофиты различаются но форме подставок: подставка женского гаметофита напоминает но форме цветок ромашки, а мужского — диск со слегка волнистыми краями. На женском гаметофите формируются женские половые органы — архегонии, в которых образуются женские гаметы — яйцеклетки. На мужском гаметофите формируются мужские половые органы — антеридии, в них образуются подвижные мужские гаметы — сперматозоиды. Как женская, так и мужская гаметы возникают в процессе митоза из гаплоидных тканей гаметофитов (n)и поэтому тоже гаплоидны (n).
Половые органы всех наземных растений многоклеточные. Многоклеточные антеридии маршанции, напоминающие длинные овальные мешки, отделяют друг от друга стерильные нити — парафизы (рис. 1).
В многоклеточных архегониях различают ножку, брюшко и шейку. Незрелую яйцеклетку защищают шейковые канальцевые клетки, разрушающиеся при созревании яйцеклетки, освобождая проход, по которому к яйцеклетке подплывают двужгутиковые сперматозоиды.
С мужского растения на женское сперматозоиды переносятся с каплями дождя или росы, после чего плывут по поверхностной пленке воды, пока нс достигнут архегония с яйцеклеткой.
При слиянии сперматозоида с яйцеклеткой образуется диплоидная зигота (2n), из которой вырастает диплоидное поколение маршанции — спорофит (2n). Спорофит представляет собой маленькую коробочку на ножке, внешне напоминающую бубенчик. Он не способен питаться самостоятельно и паразитирует на гаметофите, прикрепляясь к нему с помощью особого органа — гаустории, или стопы.
Диплоидные клетки (2n) спорогенной ткани коробочки спорофита делятся путем мейоза, образуя гаплоидные споры (n).
По окончании созревания спор коробочка раскрывается четырьмя створками и споры высыпаются на землю. Рассеиванию спор помогают специальные упругие нити — элатеры. В сухую погоду элатеры скручены, а во влажную — распрямляются, разрыхляя массу спор.
Таким образом, для всех моховидных характерно:
Чередование диплоидного поколения — спорофита (2n) и гаплоидного поколения — гаметофита (n).
В жизненном цикле моховидных доминирует гаплоидное поколение — гаметофит (n); именно он осуществляет основную функцию растения — фотосинтез.
Спорофит мхов не способен питаться самостоятельно и паратизирует на гаметофите (исключением являются антоцеротовые).
Для размножения моховидным нужна капельно-жидкая влага, поскольку мужские гаметы моховидных — сперматозоиды передвигаются в воде.
Жизненный цикл зеленого мха кукушкин лен
Жизненный цикл кукушкина льна (Polytrichum commune) очень похож на жизненный цикл маршанции, однако спорофит кукушкина льна крупнее и представляет собой коробочку на ножке.
В период созревания спор коробочка прикрыта колпачком — калиптрой. Сама коробочка устроена довольно сложно (рис. 2). В центре нее проходит вертикальная ось — колонка. Вокруг колонки на специальных нитях подвешен цилиндрический спорангий. По окончании созревания спор крышечка коробочки — оперкулум откидывается. Под крышечкой находятся пленка — эпифрагма.
Зубцы по краю коробочки (их называют перистомом) в сухую погоду отгибаются, открывая отверстия в эпифрагме, через которые высыпаются споры. Во влажную погоду зубцы перистома закрывают отверстия эпифрагмы. Наиболее существенной особенностью развития листостебельных мхов (к которым принадлежит и кукушкин лен), отличающей их от большинства печеночников, является то, что у них из споры вырастает не гаметофит, а многоклеточная зеленая нить, напоминающая зеленую нитчатую водоросль, — протонема. На протонеме формируются многоклеточные почки и уже из почки вырастает соответствующий гаметофит (рис. 3). У сфагновых и андреевых мхов, в отличие от зеленых, протонема не нитчатая, а пластинчатая.
Интересно, что у некоторых видов зеленых мхов протонема может стать основной жизненной формой.
Помимо полового размножения мхи могут размножаться и вегетативно — кусочками слоевища (таллома) или стебля с листьями. Особенно широко распространено и разнообразно но форме вегетативное размножение у печеночников. Они образуют множество органов вегетативного размножения: выводковые тельца, выводковые почки (рис. 4), придаточные побеги, ломкие листья, клубеньки и т.п.
Широко распространено вегетативное размножение и у листостебельных мхов. Это размножение частями гаметофита: разрастание моховой дернины, размножение ломкими стеблями, укороченными выводковыми ветвями, выводковыми почками, ломкими листьями, первичной протонемой. Кроме того, листостебельные мхи могут производить и специальные выводковые тельца, развивающиеся на различных частях растения, чаще всего на стеблях.
Важнейшие особенности моховидных
Для растений, принадлежащих к отделу моховидные, так же как для всех остальных наземных растений, характерно чередование гаплоидного (n) и диплоидною (2n) поколений (гаметофита и спорофита), по в отличие от сосудистых растений, у моховидных доминирует гаплоидное поколение — гаметофит (n). Диплоидное поколение — спорофит (2n), как правило, паразитирует на гаметофите (исключение составляют только антооцеротовые).
У моховидных отсутствуют или плохо развиты многие органы и ткани, возникающие у растений в связи с приспособлением к наземному образу жизни: проводящая, механическая, покровная ткани; у них нет корней (их заменяют ризоиды). Так же как водоросли, мхи впитывают воду всей поверхностью тела. Для размножения им необходима капельно-жидкая влага (их мужские гаметы — сперматозоиды передвигаются в воде).
Моховидные живут в местах с повышенной влажностью (леса, болота). Они доминируют в арктических и горных областях. Некоторые виды приспособились к обитанию на раскаленных скалах и в пустынях.
Важную экологическую роль в природе играют торфяные болота, в которых берет свои истоки большинство европейских рек.
Симбиоз мхов и насекомых
Как известно, устойчивые симбиотические отношения с насекомыми — характерная особенность цветковых растений, однако существуют мхи, привлекающие насекомых к распространению спор. Это зеленые мхи рода сплахнум (Splachmun). У представителей этого рода сильно расширено кольцо на шейке коробочки апофиза, вследствие чего вся коробочка напоминает зонтик. Зонтики крупные, до 2 см в диаметре, и ярко окрашены — красные, желтые, пурпурные. Распространителями липких спор сплахнума являются мухи, которых привлекает не только яркая окраска апофизы, но и ее запах.
На фото — маршанция изменчивая (Marchantia polymorpha), разросшаяся почти сплошным ковром на гари спустя несколько лет после лесного пожара. Это очень распространенный вид-космополит, заселяющий нарушенные местообитания вроде пожарищ или кострищ, обочин и выбоин дорог, любых обнажений почвы. Для успешного развития маршанции нужно одновременно и достаточно высокое содержание минеральных веществ в субстрате, и низкое число конкурентов. Растения с такой экологической стратегией называются рудералами.
Маршанция относится к печеночникам, одному из отделов мохообразных растений. По ряду морфологических и даже цитологических признаков печеночники заметно отличаются от двух других отделов — антоцеровых мхов (см. картинку дня «Из жизни антоцероса») и настоящих мхов.
Как вы видите, выглядит маршанция довольно замысловато. Но при этом у нее нет разделения тела на органы. Такой вариант организации тела растения называют талломом, или слоевищем. С нижней стороны таллома есть одноклеточные ризоиды. Часть из них прорастает в субстрат и, по-видимому, служит для закрепления таллома в субстарте и для минерального питания. Часть ризоидов ориентирована параллельно субстрату и прикрыта брюшными чешуйками. Эти ризоиды, отличающиеся внутренними утолщениями клеточной стенки, называют язычковыми. Есть предположение, что они работают как аналог проводящей системы и позволяют воде распространяться по слоевищу.
У всех мохообразных в жизненном цикле преобладает гаплоидная стадия — гаметофит. На нем образуются половые клетки, а после оплодотворения там же, на гаметофите, развивается диплоидное поколение — спорофит. У каждого гаметофита есть пол, мужской или женский, который определяется половой хромосомой, Y или X. Выросты, похожие на миниатюрные пальмы, — это так называемые подставки. На подставках образуются мужские гаметангии — антеридии, и женские — архегонии. В антеридиях развиваются сперматозоиды, в архегониях — яйцеклетки. Отличить мужскую подставку от женской легко: мужская подставка более плоская, более округлая (на верхнем фото мужские подставки имеют фиолетовый оттенок). Антеридии расположены с ее верхней стороны. Выход сперматозоидов происходит при попадании воды на мужскую подставку.
Скрещивание маршанции изменчивой в лабораторных условиях. На мужские подставки капают жидкость, антеридии открываются, и из них выходит мутная масса, содержащая сперматозоиды
Женская подставка разделена на 9–12 узких долей, загибающихся вниз. Архегонии закладываются сверху, но в ходе роста и растяжения клеток оказываются с нижней стороны, на которой развиваются многочисленные чешуйки. Женская подставка такой формы неплохо удерживает каплю воды. На ножках подставок есть ризоиды: ножка по строению соответствует нижней стороне слоевища. Для оплодотворения маршанции нужна пленка воды, по которой сперматозоиды могут доплыть до архегониев с яйцеклеткой. Вероятно, ризоиды и чешуйки с нижней стороны женской подставки способствуют образованию и сохранению этой водной пленки.
После оплодотворения на нижней стороне женских подставок развивается диплоидное поколение — спорофит. Спорофит состоит из коробочки, ножки и стопы — гаусториальных клеток, которые обеспечивают поглощение питательных веществ из тканей гаметофита. В коробочке происходит образование спор и специальных разрыхляющих споровую массу клеток — элатер (см. также Elater). Ножка спорофита у маршанции очень короткая, и коробочка долгое время остается скрытой в тканях материнского организма. При созревании коробочки ножка удлиняется, коробочки оказываются снаружи, раскрываются и рассеивают гаплоидные споры. Одна подставка маршанции производит около 7 миллионов спор.
Спорангии маршанции изменчивой. А–С — развитие спорангия, разные стадии. D–Е — зрелый спорангий. F — женская подставка с закрытыми зрелыми спорангиями. G — открытый спорангий со споровой массой и элатерами. H — женская подставка с открытыми спорангиями. Фото с сайта thenode.biologists.com
Прорастание спор светозависимое и по крайней мере в лабораторных условиях требует длинного светового дня. Споры разбухают и начинают синтезировать хлорофилл, потом прорастают. Первые этапы развития известны с точностью до клетки: сначала образуется первый ризоид, потом сферическая протонема из небольшого числа клеток.
Таллом маршанции образует особые структуры для вегетативного размножения — круглые корзиночки с выводковыми тельцами. Корзиночки появляются за счет удлинения клеток эпидермы. После этого другие клетки эпидермы на дне корзиночки начинают делиться и образуют выводковые тельца — маленькие многоклеточные образования с одной или несколькими точками роста. Выводковые тельца разлетаются с каплями дождя, когда капли попадают в корзиночку.
У маршанции изменчивой подставки и корзиночки с выводковыми тельцами могут встречаться на одном талломе в одно и то же время. В лабораторных условиях можно стимулировать появление подставок длинным световым днем и дальним красным светом, а образование корзиночек — коротким световым днем, то есть, как этого и следует ожидать, фазы бесполого и полового размножения запускаются разными факторами среды. Что приводит к одновременному появлению подставок и корзиночек в природе, пока не установлено.
В талломе маршанции есть несколько типов тканей: покровная ткань — эпидерма; нефотосинтезирующая основная ткань, некоторые клетки которой несут запасы питательных веществ — зерна крахмала или масляные тельца; и способная к фотосинтезу ассимиляционная ткань. Масляные тельца, образующиеся из фрагментов эндоплазматической сети, — это уникальная органелла, известная только у печеночников. В них содержится смесь гидрофобных веществ разного состава, в основном терпеноиды, а также ароматические вещества, ненасыщенные жирные кислоты и триглицериды. Некоторые из этих веществ в экспериментах проявляют цитотоксичность по отношению к клеткам животных, антибактериальную и антигрибную активность. В масляных тельцах маршанции также обнаружены белки, родственные ферментам синтеза изопренов. Значение масляных телец точно не установлено, но можно предположить, что эти вещества могут отпугивать животных, желающих полакомиться сочным талломом, защищать талломы от ультрафиолетового излучения, подавлять развитие патогенных грибов.
Срез слоевища маршанции. А — воздушная камера, В — брюшные чешуи, С — ризоиды. Ассимиляционная ткань окрашена фиолетовым, обратите внимание, какая она тонкая. Срез прошел поперек язычковых ризоидов, пучки которых идут под защитой брюшных чешуй. Длина масштабного отрезка — 0,2 мм. Изображение с сайта commons.wikimedia.org
Ассимиляционная ткань образует воздушные камеры, которые сообщаются с атмосферой через поры, одна пора на одну камеру. Они способствуют газообмену и благодаря водоотталкивающей кутикуле не позволяют каплям воды проникнуть в воздушные камеры. Иногда эти поры называют устьицами, но это не совсем корректно: несмотря на общую функцию, строение пор маршанции совсем непохоже на строение устьиц мхов из других отделов и на устьица сосудистых растений. Эти поры, за редкими исключениями, не способны менять свой диаметр при обезвоживании таллома (исключение — печеночники рода Preissia с особым строением пор). Удивительно и то, что они образуются у гаплоидного поколения, у всех остальных высших растений устьица есть только на спорофитах.
Схема среза воздушной камеры маршанции. Бочонкообразная структура — пора. Она состоит из 4–7 расположенных друг над другом колец, каждое из 4–5 клеток. Рисунок с сайта elte.prompt.hu
Хотя у маршанции нет корней, она может образовывать микоризоподобные симбиозы с гломусовыми грибами. Контакт клеток гриба и печеночника происходит внутри таллома. Для маршанции изменчивой образование такого симбиоза не очень характерно и отмечено только для одного подвида — Marchantia polymorpha subsp. montivagans, который растет на более бедной почве, чем остальные подвиды. Несколько видов грибов-базидиомицетов специализировались на питании маршанцией. Эти грибы относят к роду Loreleia. Точно не установлено, являются ли они сапротрофами или паразитами.
Рудеральные растения, к которым относится маршанция, обычно легко культивировать: воссоздание нарушенных местообитаний требует минимум усилий. И как раз рудеральные виды зачастую становятся модельными объектами биологов. Арабидопсис, или резуховидка Таля, знаменитое цветковое растение, на котором изучены основные закономерности физиологии растений, — сорняк, как и маршанция. Первые лабораторные эксперименты с маршанцией датируются XIX веком. На ней проследили развитие выводковых телец и оплодотворение. В эпоху молекулярно-генетических исследований маршанция изменчивая оказалась одним из первых растений, у которых отсеквенировали геном митохондрий и пластид. Для маршанции разработаны протоколы выращивания и генетической трансформации. Недавно была установлена и проанализирована полная последовательность генома маршанции изменчивой.
Мотивация изучать маршанцию не ограничивается удобством ее выращивания. Исследователей интересует ее сложное и своеобразное строение, и они сопоставляют закономерности развития разных структур в гаметофитах маршанции и спорофитах сосудистых растений. Геном маршанции сравнивают с геномами высших растений, пытаясь понять положение печеночников на эволюционном древе высших растений. Есть серьезные аргументы как в пользу того, что печеночники представляют собой базальную группу по отношению ко всем остальным высшим растениям, так и в пользу того, что это сестринская группа к настоящим мхам. Если печеночники — базальная группа, то можно предположить, что в жизненном цикле последнего общего предка всех высших растений тоже преобладала гаплоидная стадия, и в таком случае резонно звучит гипотеза сдвига экспрессии регуляторных генов (см.: Современные высшие растения возникли в результате сдвига экспрессии генов, «Элементы», 10.04.2018). Если же все три отдела мохообразных объединяются в монофилетическую группу, сестринскую по отношению к сосудистым растениям, то мы не можем ничего утверждать о жизненном цикле последнего общего предка всех высших растений. Но в любом случае, изучение печеночников и сравнение их с другими высшими растениями позволяет что-то узнать о путях эволюции и о том, как мог выглядеть и функционировать загадочный предок высших растений.